PiRNA
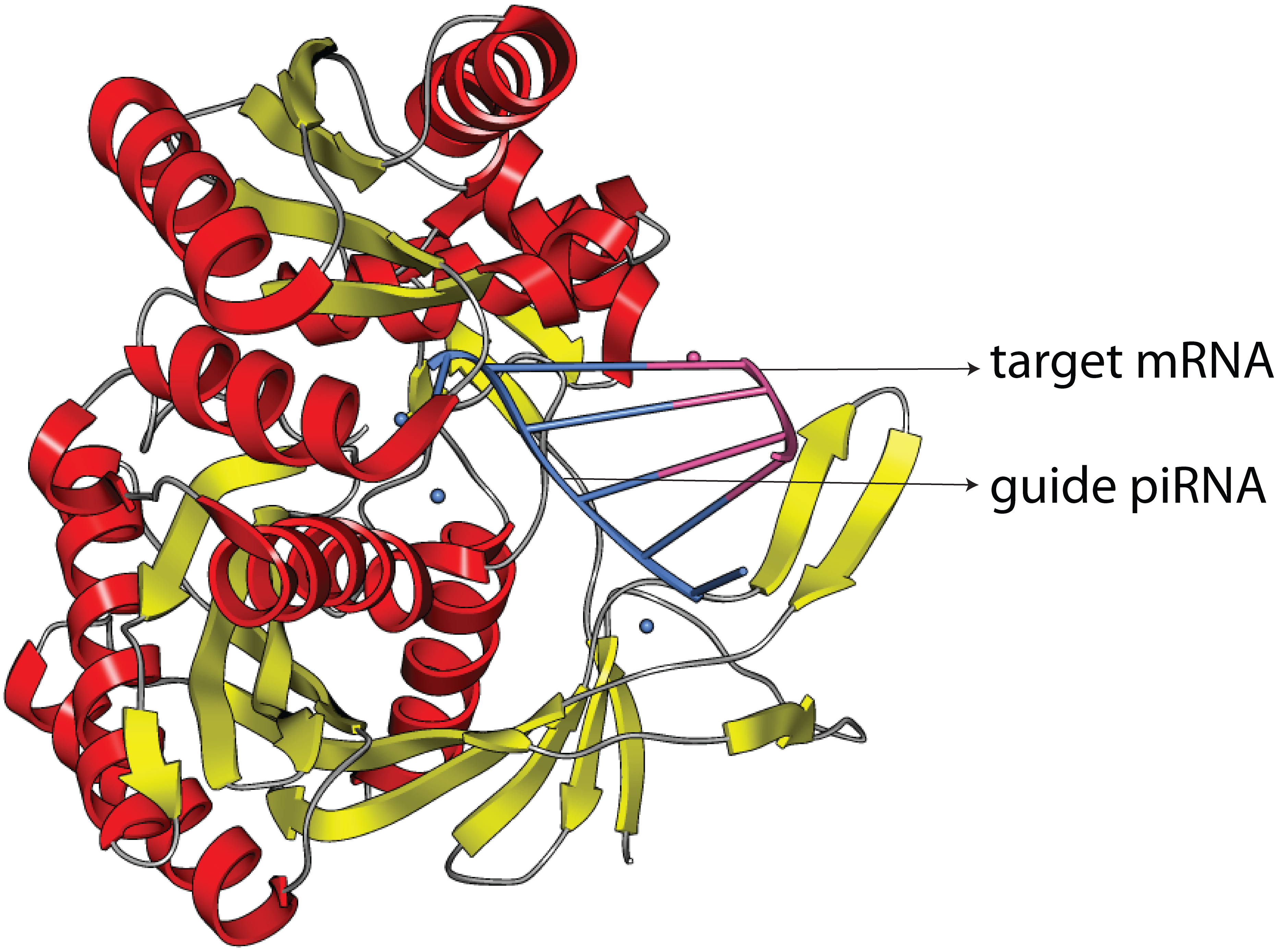
Piwi-interacting RNA (piRNA)是一大类主要在动物体内表达的small non-coding RNA,piRNA通过与Piwi蛋白互作形成RNA-protein复合体。该piRNA复合体在生殖细胞中参与表观遗传和逆转录转座子(retrotransposons)的转录后基因沉默。piRNA与miRNA和siRNA在长度、序列结构和生物起源上均存在差异;
PiRNA特点
1)26~31nt
2)无明显二级结构
3)5’端第一个碱基为U
4)5’端单磷酸盐(monophosphate)和3’端修饰(2’-O-methylation modification)阻止2’ or 3’氧化,增加PiRNA稳定性。
5)种类较多,不具一定的保守性,老鼠体内 50,000 unique piRNA,果蝇中>13,000。
6)产生存在显著的链偏好性;
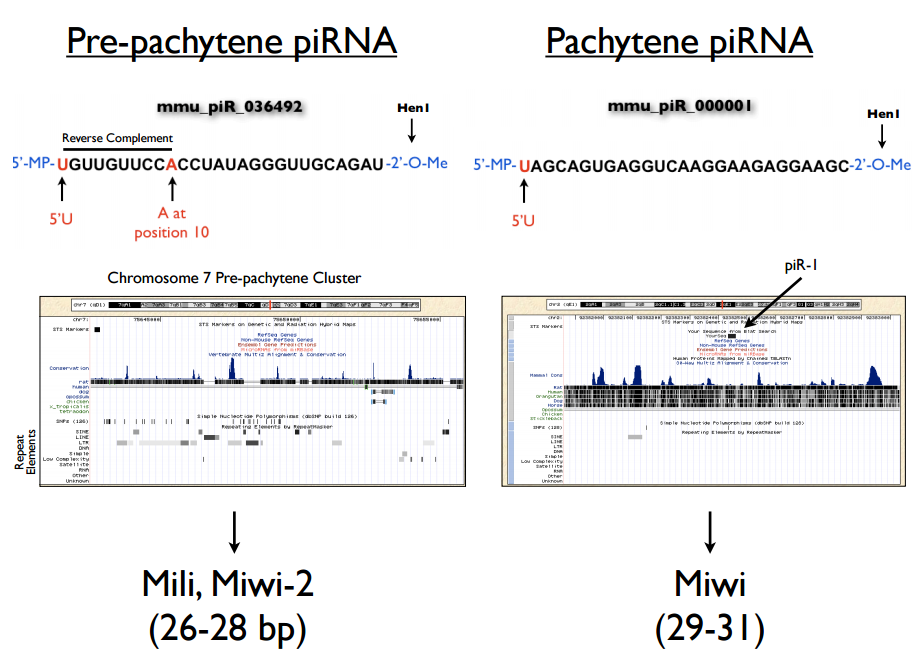
位置
成簇贯穿基因组中,其每一个簇中包含PiRNA小于10个或达到成千上万,且大小差异极大。
在果蝇和脊椎动物中定位于非编码基因间,在线虫蛋白编码基因间也鉴定到PiRNA。
在无脊椎动物和哺乳动物生殖细胞中较多。
细胞核和细胞质中均存在。
生物起源
PiRNA的产生存在显著的链特异性,可能仅仅是来源于双链DNA的某一条链,这表明转录的长的单链前体经过一次初加工形成 pachytene PiRNA,这过程中PiRNA前体的转录趋向于起始于5’端第一个碱基U。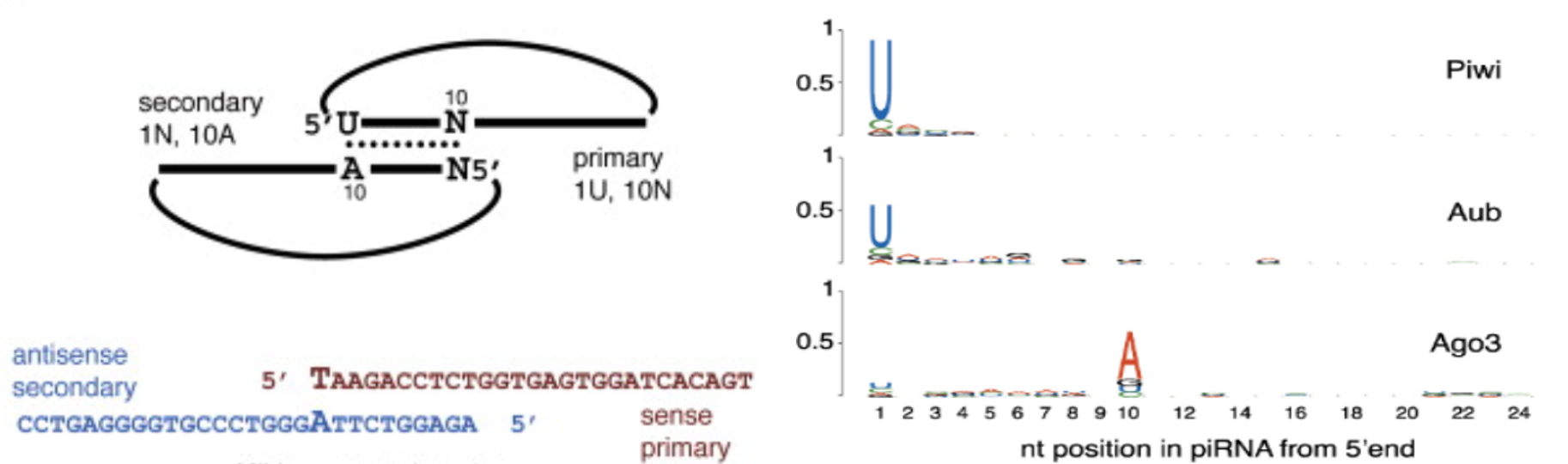
‘Ping Pong’机制:初级PiRNA(Primary piRNAs)第一个碱基位置偏向为U,第10个无偏向性;次级PiRNA(Secondary piRNAs)(产生于初级PiRNA指导的剪切)第一个碱基无偏向,第10个偏向A;
二者从5’端开始有10个碱基的互补。More:A piRNA Pathway Primed by Individual Transposons Is Linked to De Novo DNA Methylation in Mice
初级PiRNA识别其互补靶标并招募Piwi蛋白,然后从距离初级PiRNA 5’端10个碱基处劈开(识别的互补靶标)形成次级PiRNA,次级PiRNA靶向到第10个碱基是 A 。
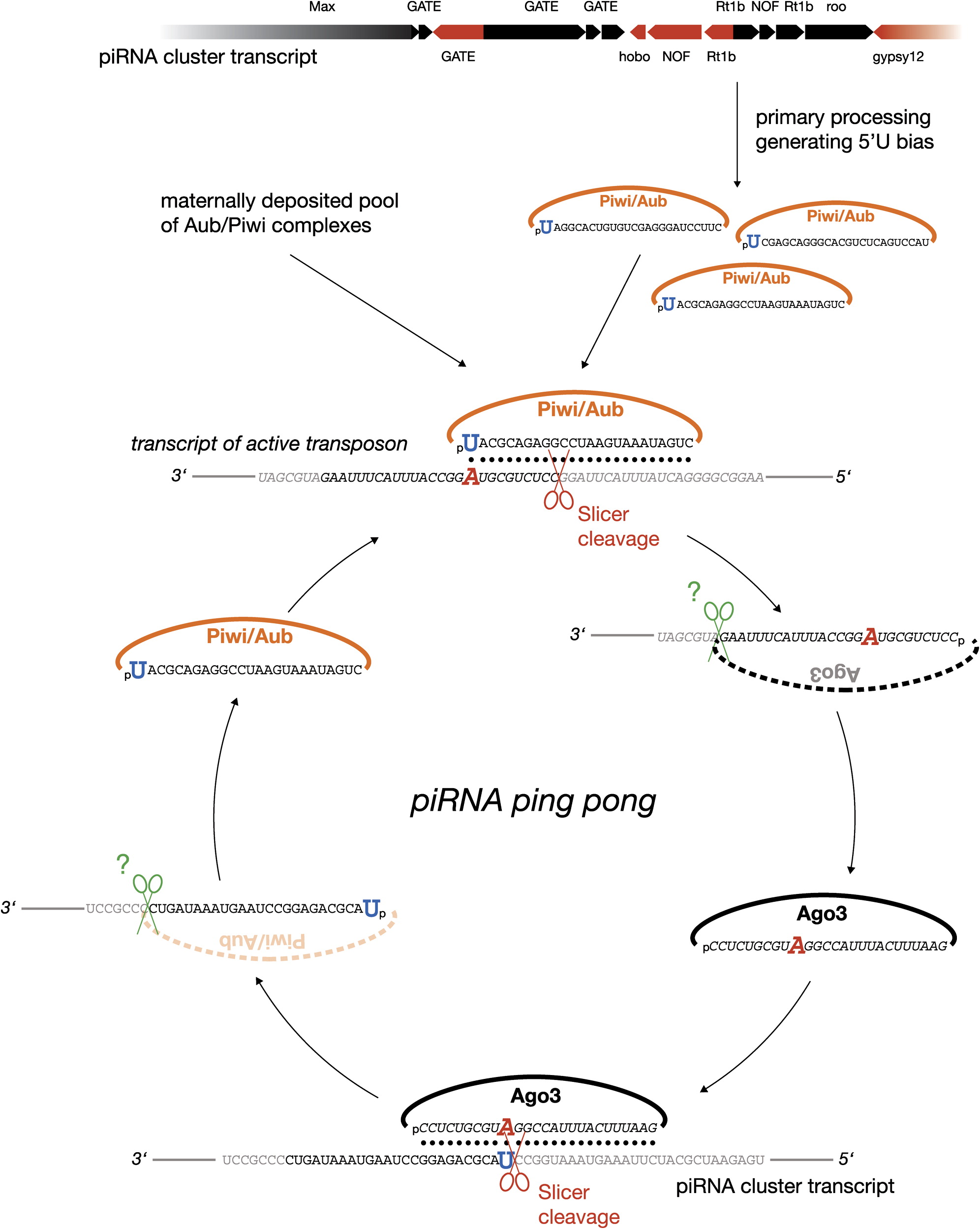
Discrete Small RNA-Generating Loci as Master Regulators of Transposon Activity in Drosophila
生物功能
沉默转座子
见More:Piwi蛋白:RNAi中作用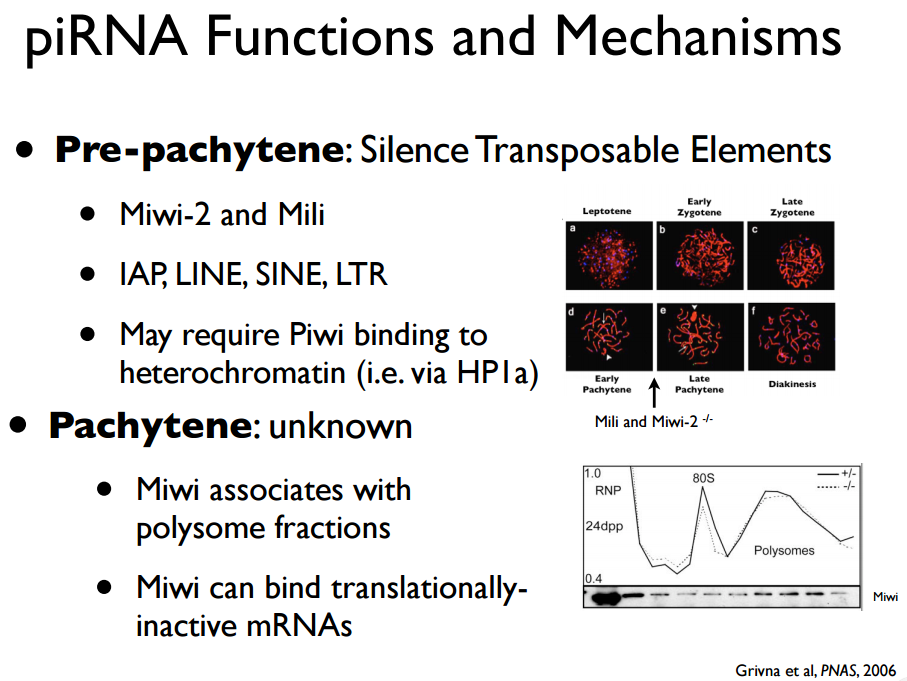
后生效应(Epigenetic effects)
动植物中,小RNA通过特定的胞嘧啶甲基化来间接调控表观遗传,并且小RNA自身承担着表观遗传信息的载体。存在某些特殊转座子差异的果蝇品系间杂交能引起后代不育,这称之为杂种不育。当这一转座子是父系遗传时不育表型表现为显性,而母系遗传能够维持育性。在P- and I-element-mediated杂种不育中,依赖于父母本不同,其作用于每一个靶标元件(element)的PiRNA数量在后代表现出明显差异,这种差异来源于受精作用。综上表明母本生殖细胞内的PiRNA对上述特殊转座子的沉默响应起到重要作用,此沉默效应的缺失将引起杂种不育。
More:An epigenetic role for maternally inherited piRNAs in transposon silencing
PiRNA鉴定
PiRNA的鉴定目前主要通过识别’ping pong‘标签,相关软件如下:
piRNABank: a web resource on classified and clustered Piwi-interacting RNAs
PingPongPro:a software for finding ping-pong signatures and ping-pong cycle activity
proTRAC: a software for probabilistic piRNA cluster detection, visualization and analysis
piRNA cluster: database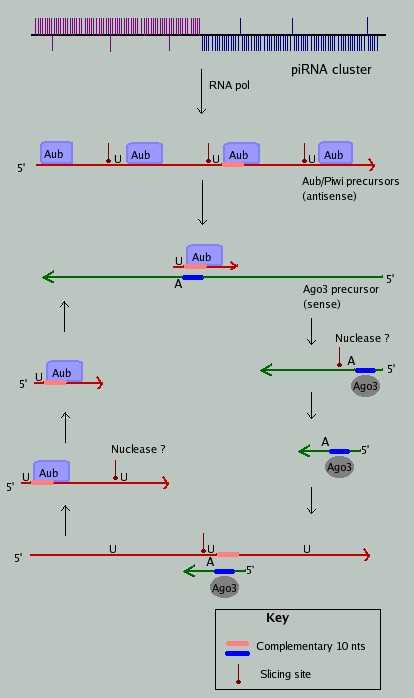
PiRNA起源
基因组重复区域,例如逆转录转座子区;
异染色质区,双链RNA的反义链;
Argonaute蛋白家族
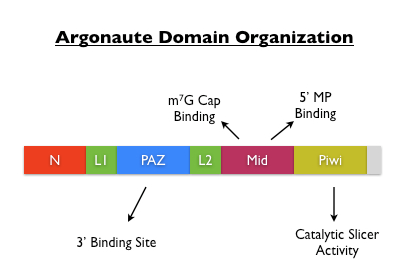
Argonaute蛋白包含有N-terminal, PAZ (Piwi-Argonaute-Zwille), middle and the C-terminal PIWI (P-element-induced wimpy testis) domains (Tolia et al., 2007)。
在果蝇中存在5种类型的Argonaute蛋白:AGO1, AGO2, Aubergine (Aub), Piwi and AGO3 (Gunawardane et al., 2007);
AGO1和AGO2属于Argonaute (AGO)亚家族,Aub, Piwi and AGO3多存在于生殖细胞系中,且属于PIWI亚家族;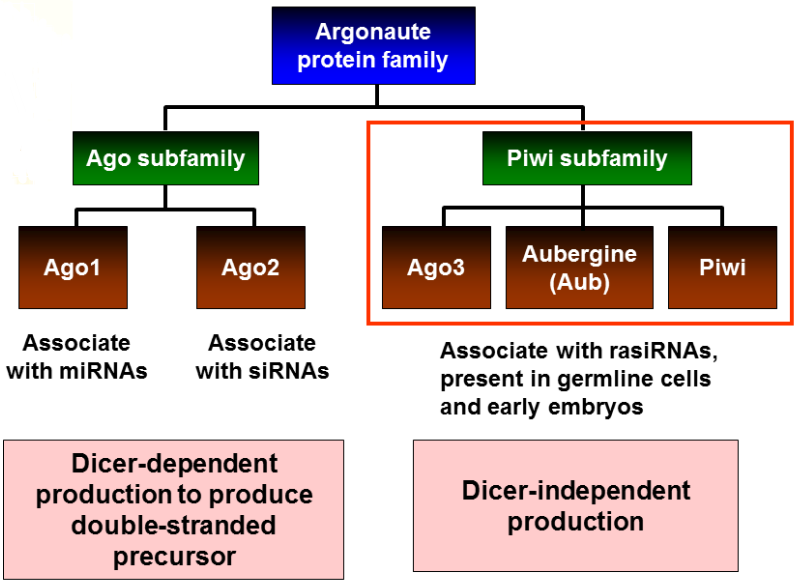
Piwi蛋白
Piwi蛋白(最初在果蝇中的P-element induced wimpy testis),维持干细胞的不完全分化和生殖细胞细胞分裂比率的稳定性。Piwi蛋白高度保守,广泛存在于动植物体内。
RNAi中作用
Piwi蛋白存在有PAZ domain,该domain在Argonaute蛋白家族中参与双链RNA导向的单链RNA的水解作用。Argonaute是广泛研究的核酸结合蛋白(nucleic-acid binding)家族,其本质上是一种RNase H-like酶,完成RNA-induced silencing complex (RISC)的催化功能。在细胞RNAi反应中,RISC复合体中的Argonaute蛋白能够绑定(bind)到由ribonuclease Dicer切割(Dicer-2)外源双链RNA的正义链和反义链产生的siRNA(small interfering RNA)和切割(Dicer-1)内源非编码RNA(non-coding RNA)产生的miRNA(microRNA)上,从而形成RNA-RISC complex。该RNA-RISC complex绑定和切开与RNA(siRNA或miRNA)碱基互补的mRNA,破坏并且阻止其翻译过程。
补充RNAi中的RdRP机制:在线虫的研究中发现, siRNA 是合成 dsRNA 的特殊引物, 在RNA 依赖RNA 聚合酶(RdRP)作用下, 以靶mRNA 为模板合成dsRNA 。新生成的dsRNA 在Dicer 酶的作用下, 裂解产生新的siRNA , 新生成的siRNA 又可进入上述循环。大量集中的siRNA 可以形成RISC复合物, 这样可以提高mRNA 降解的效率。在这种RNAi过程中, 对靶mRNA 的特异性扩增有助于增强RNAi的特异性基因监视功能, 每个细胞只需少量的dsRNA就能完全关闭相应基因的表达,该模型称为RdRP。
Piwi蛋白和转座子沉默
Piwi蛋白通过与PiRNA形成内源系统来沉默内源自私基因(endogenous selfish genetic elements)表达,例如逆转录转座子和重复序列,防止该自私基因产物干扰生殖细胞的形成。
selfish genetic elements明显特征:通过形成额外拷贝数在基因组中传播(转座子)和对宿主的成功繁殖没有特殊贡献。
RasiRNA
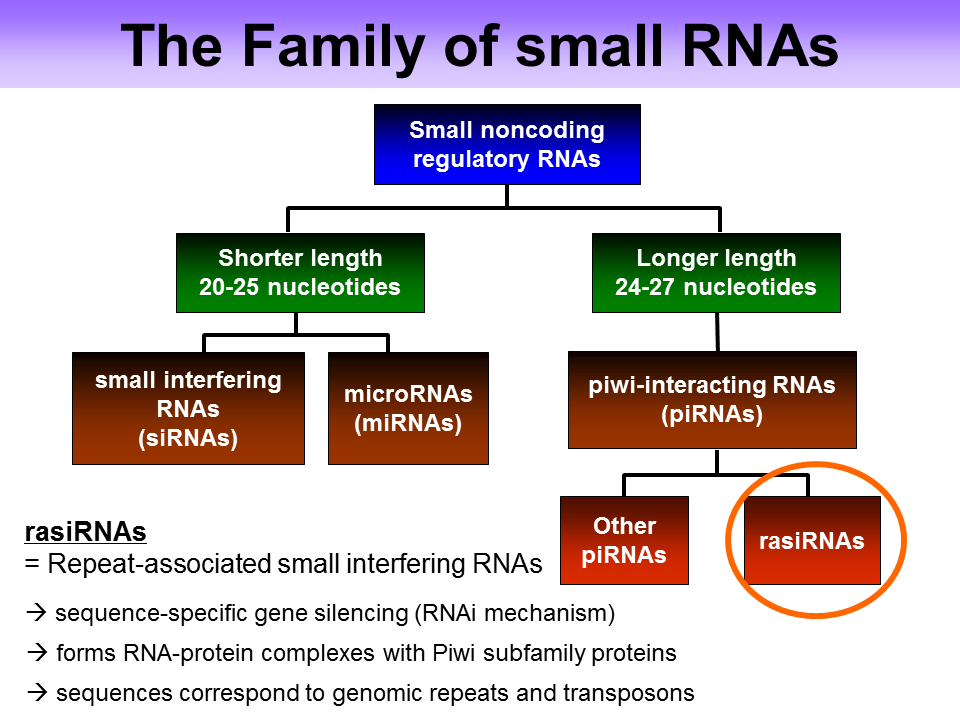
RasiRNAs(Repeat associated small interfering RNA)是piRNA的亚种,与Piwi蛋白(Argonaute蛋白家族分枝)互作参与RNAi反应。在生殖细胞中建立和维持异染色质结构,控制重复序列的转录,沉默转座子和逆转录转座子。主要产生自反义链(antisense strand),缺乏动物siRNA and miRNA所特有的2’,3’羟基末端。
More:A Distinct Small RNA Pathway Silences Selfish Genetic Elements in the Germline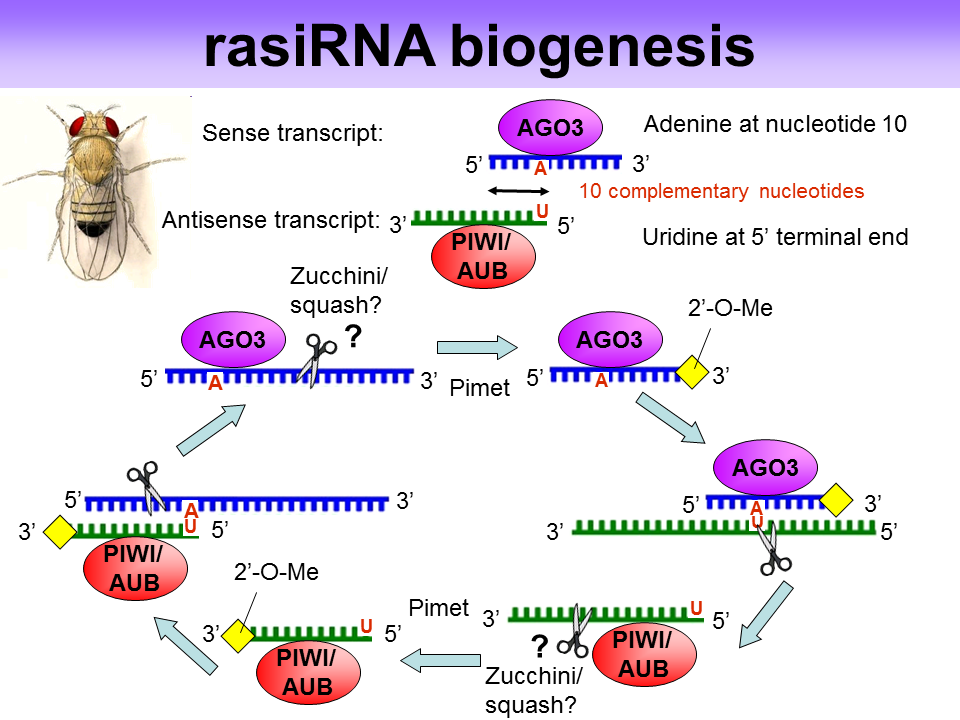
RNA 百科
RNA wiki
